 | | |
Le cytosquelette | ||
| |
Le cytosquelette est l'élément qui différe le plus les eucaryotes des procaryotes. Tous les eucaryotes ont un cytosquelette développé alors que les procaryotes n'en ont pas, à l'exception des spirochè qui possèdent des molécules semblables à de l'actine mais sans atteindre la sophistication d'un vrai cytosquelette.
Le cytosquelette est un réseau de fibres intracellulaires. Il est constitué de trois grandes familles de protéines, très conservées dans l'évolution : les filaments épais de tubuline ou microtubules, les filaments fins d'actine ou microfilaments et les filaments intermédiaires. Les deux premières familles sont très homogènes et ont été très étudiées. Elles sont présentes dans toutes les cellules et les molécules constitutives sont les mêmes, les généralités qui vont suivre s'appliquent à eux seuls. Les filaments intermédiaires sont en revanche plus hétérogène, constitués de molécules qui diffèrent selon le type cellulaire, ils ne seront pas abordés ici.
Bien que de composition et de fonction très différentes,
les filaments du cytosquelettes ont des propriétés communes.
Les microtubules et les microfilaments sont constitués d'une charpente
protéique fibrillaire formée par la polymérisation
d'une ou de deux molécules de protéine apparentée,
stabilisée par des protéines annexes appartenant à
d'autres familles.
 Ces filaments sont dynamiques (c'est pourquoi certains spécialistes
préfèrent parler de cytomuscle), les protéines constitutives
peuvent exister sous deux formes dans la cellules : monomèriques,
solubles et dispersées dans le cytoplasme ou insolubles et organisées
en filaments. Il existe un seuil de concentration des monomères au
dessus duquel les filaments se forment par polymérisation des unités
moléculaires et en dessous duquel ils se dépolimérisent.
Le filament est polarisé, une extrémité noté
+ à un seuil plus bas que l'autre extrémité noté
-. A concentration égale, le coté + croit plus vite (ou se
dépolarise - vite que le coté -). Il existe également
une plage de concentration pour laquelle le coté + croit alors que
le coté - decroit, cela entraine une migration du filament à travers
la cellule, où s'il a une position fixe, un phénomène de
tapis roulant. La polymérisation et la dépolymérisation
consomment de l'energie et la valeur des seuils est controlé par
la cellule de manière très fine.
Ces filaments sont dynamiques (c'est pourquoi certains spécialistes
préfèrent parler de cytomuscle), les protéines constitutives
peuvent exister sous deux formes dans la cellules : monomèriques,
solubles et dispersées dans le cytoplasme ou insolubles et organisées
en filaments. Il existe un seuil de concentration des monomères au
dessus duquel les filaments se forment par polymérisation des unités
moléculaires et en dessous duquel ils se dépolimérisent.
Le filament est polarisé, une extrémité noté
+ à un seuil plus bas que l'autre extrémité noté
-. A concentration égale, le coté + croit plus vite (ou se
dépolarise - vite que le coté -). Il existe également
une plage de concentration pour laquelle le coté + croit alors que
le coté - decroit, cela entraine une migration du filament à travers
la cellule, où s'il a une position fixe, un phénomène de
tapis roulant. La polymérisation et la dépolymérisation
consomment de l'energie et la valeur des seuils est controlé par
la cellule de manière très fine.
Au réseau fibrillaire d'autres types de protéines peuvent se
fixer. Certaines protéines sont fixes et assurent la liaison des
réseaux intracellulaires avec le reste de l'architecture cellulaire.
D'autre molécules sont consommatrice d'énergie appartiennent
à l'ensemble des moteurs. Elles possèdent un point d'ancrage
sur les filaments et peuvent migrer le long de ceux ci, et également
un autre point d'ancrage ce qui leur permet de transporter des protéines
ou organites le long de ce réseau. La protéine fixée
peut être un autre moteur associé à son filament, ce qui
permet d'organiser la structure du cytosquelette et déplaçant
les filaments les uns par rapports aux autres.
Les microtubules sont les plus épais (25nm) des filaments cytosqueletique. Ils interviennent dans tous les mouvements intracellulaires, migration des organites dans la cellule, transport axonal dans les neurones mais aussi séparations des deux jeux de chromosomes lors de la division cellulaire.
 Les microtubules sont des petits tubes de 25 nm de diamêtre,
la lumière faisant 10 nm. Ils sont formé par la polymérisation
de deux protéines différentes : les tubulines. La tubuline
alpha s'associe avec la tubuline beta pour former des doublets. A ce niveau,
le GTP se lie au doublet sur un site de la tubuline beta. Un changement
de conformation des protéines permet à ces doublets de s'assembler
pour former un protofilament. Enfin, 13 protofilaments forment ensemble
un filament creux de microtubule. Deux protéines différentes
peuvent se fixer à l'édifice. La dynéine, présente
notamment dans les cils et les flagelles et la protéine tau. On
peut trouver dans la cellules des microtubules doubles ou triple. Il s'agit
de microtubules qui ont en commun quelques protofilaments, 3 au niveau
de chaque point de jonction.
Les microtubules sont des petits tubes de 25 nm de diamêtre,
la lumière faisant 10 nm. Ils sont formé par la polymérisation
de deux protéines différentes : les tubulines. La tubuline
alpha s'associe avec la tubuline beta pour former des doublets. A ce niveau,
le GTP se lie au doublet sur un site de la tubuline beta. Un changement
de conformation des protéines permet à ces doublets de s'assembler
pour former un protofilament. Enfin, 13 protofilaments forment ensemble
un filament creux de microtubule. Deux protéines différentes
peuvent se fixer à l'édifice. La dynéine, présente
notamment dans les cils et les flagelles et la protéine tau. On
peut trouver dans la cellules des microtubules doubles ou triple. Il s'agit
de microtubules qui ont en commun quelques protofilaments, 3 au niveau
de chaque point de jonction.
En réponse à un signal cellulaire, le GTP peut s'hydrolyser pour former du GDP. Le microtubule devient alors instable. Il peut se dépolymeriser totalement en quelques seconde, ce qui est beacoup plus rapide que ce que permettrait une dépolarisation par les extrémité comme evoquée dans l'introduction. La polymérisation et la dépolymérisation des microtubules est donc un phénomène dynamique, finement contrôlées par la cellule.
In vitro, la polymérisation nécessite la présence
pour démarrer de centre de nucléation, c'est à dire
de très petits fragments de microtubules. Dans la cellule, de tels
centre n'existenr, mais une structure en tiens lieu, le centre organiteur
des microtubules (COMT). Chaque cellule possède un ou plusieurs
COMT. Ils sont constitués d'un centrosome entouré d'un ensemble
complexe de protéine encore mal identifiées avec des granules
denses
aux électrons à la périphérie de la structure.
Le centrosome est constitué de deux cylindres de 9 triplets de microtubules
(les centrioles) positionnés à angle droit. Ce centrosome
est doué de continuité génétique, bien qu'aucun
ADN n'ait été trouvé dans la structure, les deux centrioles
se séparent, chaque centriole isolé déclenche la synthèse
d'un autre centriole pour reconstituer un centrosome complet. Un centrosome
ne peut pas apparaître à partir de rien dans une cellule.
Cette propriété laisse penser que le centrosome et le flagelle
(seul organite à base de centrosome) serait tout ce qui resterait
d'une endosymbiose. Toutefois, comme aucun procaryote actuel ne possède
de centrosome, l'origine de l'organisme serait donc dans un type nouveau
(ni bactérie, ni archéobactérie) qui aurait aujourd'hui
disparu ou n'aurait pas encore été découvert. D'autres
scientifiques situent toutefois son origine dans les spirochète, la
question est encore débattue.
La présence d'un COMT modifie les propriété des
microtubules. L'extrémité - est à proximité
du COMT alors que la + est est éloignée. La concentration
nécessaire pour obtenir une polymérisation est très
abaissée, et une baisse de la concentration en tubuline ne diminue
pas la longueur des filaments mais leur nombre. Tout ce passe comme si
le COMT avait plusieurs sites de nucléation dont le nombre dépend
de la concentration en tubuline, tout centre inoccupé donne aussitôt
naissance à un nouveau filament.
En dehors des microtubules qui traversent la cellule, ils participent à
la formation de structures plus complexes. Les deux que nous verrons sont
le fuseau mitotique et les flagelles.
Ce fuseau de microtubule intervient lors de la mitose dans la séparation des deux jeux de chromosomes entre les cellules filles. Avant la mitose, toutes les structures microtubulaires de la cellule se dépolymèrisent, cils et flagelles compris. Au départ, l'aster, situé à proximité du noyau, se divise, chaque aster se déplace à une extrémité de la cellule, filant derrière eux le fuseau mitotique. Chaque aster est en réalité un COMT, mais sont aspect en étoile viens de ce que tous les centres de nucléation doivent être occupés, pas seulement ceux impliqués dans la formation du fuseau. Les autres filaments rayonnants sont les fibres astrales. Les filaments ne sont pas continuité d'un aster à l'autre, la cohérence est assurée au centre par des protéines connectrices.
Les filaments du fuseau mitotique vont se fixer par leur extrémité + à une structure particulière du centromère des chromosomes condensés, le kinétochore. Chaque kinétochore reçoie une quarantaine de filaments. Quand tous les centromères sont fixés à des microtubules, les deux chromatides de chaque chromosome se séparent. Les microtubules vont alors se retracter, entrainant les chromatides avec elles. En fait, c'est le chromatide qui migre le long du microtubule grace à des molécules de kinésines présentent dans le kinétochorere, le filament se dépolarisant juste derrière lui.
A la fin de la mitose, les deux jeux de chromosomes sont donc séparés, chacun à une extrémité de la cellules. Le fuseau mitotique va se désorganiser, la tubuline servira à reconstruire le cytosquelette des deux nouvelles cellules. Après la séparation effectives des cytoplasme, la cellule va reconstituer toutes ses structures microtubulaire.
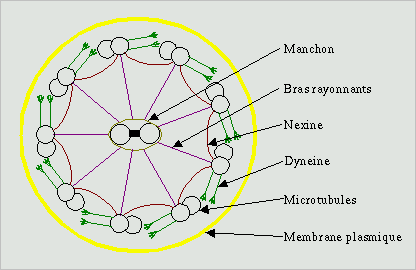 Les flagelles et les cils sont des expansions membranaires extracellulaires
qui possèdent la propriété de battre. La différence
entre les deux structures est la taille qui conditionne le mode de fonctionnement
: le flagelle est plus long que la longueur d'onde du battement, il ondule;
le cil est plus court, il bat. La forme du flagelle est assurés
par une charpente de microtubules, l'axonème (image ci contre), au coeur de l'expansion
membranaire. Le centre est occupé par un doublet de microtubules
enveloppé d'un manchon protéique. Ce doublet est entouré
d'un cylindre de 9 doublets de microtubules partiellement fusionnés. Ces doublets sont reliés
entre eux par des bras de dynéine et avec le doublet central par
les bras rayonnants. À la base du flagelle, dans le cytoplasme, se
trouve le corps basal. Il est constitué de 9 triplets de microtubules
disposés en cylindre. De microtubules de chaque triplet sont en
continuité avec ceux des doublets periphériques de l'axonème,
le doublet central s'arrête à la limite du cytoplasme et n'arrive
pas au corps basal.
Les flagelles et les cils sont des expansions membranaires extracellulaires
qui possèdent la propriété de battre. La différence
entre les deux structures est la taille qui conditionne le mode de fonctionnement
: le flagelle est plus long que la longueur d'onde du battement, il ondule;
le cil est plus court, il bat. La forme du flagelle est assurés
par une charpente de microtubules, l'axonème (image ci contre), au coeur de l'expansion
membranaire. Le centre est occupé par un doublet de microtubules
enveloppé d'un manchon protéique. Ce doublet est entouré
d'un cylindre de 9 doublets de microtubules partiellement fusionnés. Ces doublets sont reliés
entre eux par des bras de dynéine et avec le doublet central par
les bras rayonnants. À la base du flagelle, dans le cytoplasme, se
trouve le corps basal. Il est constitué de 9 triplets de microtubules
disposés en cylindre. De microtubules de chaque triplet sont en
continuité avec ceux des doublets periphériques de l'axonème,
le doublet central s'arrête à la limite du cytoplasme et n'arrive
pas au corps basal.
Les flagelles battent par glissement des doublets de microtubules entre eux. C'est la dynéine, qui en hydrolysant l'ATP, assure ce glissement. L'axonème étant cylindrique et le glissement se produisant dans le même sens relatif pour tous les doublets, la structure devrait se vriller. Ce sont les autres proétines qui transforment ce vrillage en battement.
Les microtubules sont impliqués dans la répartition des éléments intracytoplasmiques. Ils sont responsables d'une part de l'intégrité des structures cellulaires, d'autre part des mouvements intracytoplasmiques, comme montré dans le cas du fuseau mitotique. Les microtubules sont associé à un moteur protéique constitué de kinésine. Ce moteur utilise les microtubules comme rails pour déplacer des organites ou d'autres microtubules. Les microtubules constituent donc le système majeur de répartition intracellulaire.
Les microfilaments sont des filaments fins (9 nm) constitué d'actine, éventuellement ramifiés. Ils sont impliqués dans des mouvements de grande ampleur impliquants la déformation de la structure cellulaire : contraction, migration, pseudopode, à l'exception des mouvements des cils et flagelles dont le moteur est constitué de tubuline. Leur polymérisation et leur dépolimérisation permet à la cellule de contrôler la fluidité du cytoplasme et de générer des mouvements qui permettent à la cellule de migrer. A cela s'ajoute l'effet des protéines motrices qui augmentent leurs possibilités
Les microfilaments sont constitués d'un coeur d'actine associé à diverses protéines accessoires. Le monomène d'actine est l'actine G (pour globulaire). Cette protéine est tellement conservée au cours de l'evolution que l'on peut obtenir des structures fonctionnelle en mélangeant l'actine G provenant de plusieurs espèces. La polymérisation de l'actine produit un brin en forme de double hélice. L'actine F (pour fimanent) est stabilisé par un filament de tropomyosine inséré dans le sillon large de la double hélice. Diverses protéines assurent le coiffage du filament (structure en bout de filament pour stabiliser sa longueur), le pontage de deux filaments pour construire des structures en 2 ou 3 dimensions et des protéines d'ancrage à d'autres structure.
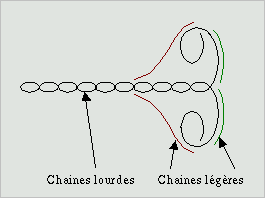 Les propriétés contractiles des filaments d'actine sont dûes
à une famille de protéines motrices spécifiques :
les myosines. Contrairement à l'actine, les myosines sont très
diversifiées. Les différences jouent sur le système
de régulation de la contraction que sur l'organisation spatiale
des filaments de myosine. Le monomère de myosine est constituée
d'une longue queue, portant deux têtes flexibles à une extrémité,
la molécule est en forme de Y. La tête flexible possède
un site de liaison à l'actine et un site pour l'ATP, c'est elle
qui est responsable de l'aspect moteur de la protéine, la queue
ayant une fonction d'ancrage.
Les propriétés contractiles des filaments d'actine sont dûes
à une famille de protéines motrices spécifiques :
les myosines. Contrairement à l'actine, les myosines sont très
diversifiées. Les différences jouent sur le système
de régulation de la contraction que sur l'organisation spatiale
des filaments de myosine. Le monomère de myosine est constituée
d'une longue queue, portant deux têtes flexibles à une extrémité,
la molécule est en forme de Y. La tête flexible possède
un site de liaison à l'actine et un site pour l'ATP, c'est elle
qui est responsable de l'aspect moteur de la protéine, la queue
ayant une fonction d'ancrage.
Bien que les modes de régulation soient très diversifiés, le fonctionnement de tous les types de myosine est similaire. En présence de calcium et d'ATP, la tête de la myosine s'accroche au filament d'actine. L'hydrolyse de l'ATP provoque la rotation de la tête et sa séparation du filament d'actine. Le départ de l'ADP de la tête de la myosine lui permet de reprendre sa forme initiale. A la fin du mouvement, la molécule de myosine a glissé le long du filament d'actine. Le sens d'accrochage du filament d'actine par rapport à la myosine est polarisé. Pour que le mouvement ait lieu, il est indispensable que les deux éléments actine et myosine soient correctement orientés.
Les molécules de myosines forment 3 types de structures en fonctions des besoins de la cellules :
Les microfilaments sont inclus dans deux grands réseaux filamenteux: un reseau sous membranaire et les cables de stress.
Le réseau sous membranaire est un réseau de filaments d'actine situé sous la membrane plasmique. Il constitue une charpente similaire à la charpente des domes géodésiques. Ce réseau permet à la cellule de contrôler sa forme et de participer au déplacememt.
Les cables de stress sont des filaments d'actine qui traversent le cytoplasme. Ils sont ancrés d'un coté à la membrane plasmique au niveau des points focaux et de l'autre à un point focal de la membrane opposé ou à une structure cytoplasmique appelé corps dense. Les points focaux sont des assemblages de protéines dont l'une d'entre elle, la taline est transmembranaires. Chez les animaux, la partie extracellulaire de ces protéines est relié à la matrice extracellulaire ce qui assurent la continuité mécanique du tissu de par et d'autre de la membrane plasmique. Le cable de stress est constitué d'une succession de corps denses composés d'alpha actinine. Ces corps denses servent d'ancrage à des filaments fins d'actine, l'extrémité + est fixée aux corps dense. L'actine ne relie pas deux corps denses, mais s'arrête au milieu d'entre eux ou se trouve l'extrémité - du filament, ce sont des rubans bipolaires de myosine qui assurent la continuité du cable. Les cables sont disposés de façon à résister aux forces qui s'exercent dans la cellule.

D'autres structures impliquent les filaments d'actine : les voiles de migration des fibroblastes, les spikes des neurites en croissance, les stéréocils des cellules auditives, etc...

