 | | |
Les organites des eucaryotes | ||
| |
Il existe deux principales différences entre les eucaryotes et les procaryotes, la présence chez les premiers d'organites intracellulaires et l'existence du cytosquelette, un réseau de protéine fibrillaire qui participe à la forme et aux mouvements de la cellule. Les organites, décrits sur cette page sont chargés d'accomplir une ou plusieurs tâches bien précises dans la cellule. En fait, l'architecture des eucaryotes n'a rien à voir avec celle des procaryotes, mais si les mêmes éléments de base se retrouvent. En revanche certains organites ressemblent tellement à des procaryotes que la majorités des scientifiques considèrent à l'origine qu'ils en étaient et que les eucaryotes sont le résultat d'une symbiose entre une cellule de type encore indeterminée (mais certainement une archéobactérie) et plusieurs bactéries. Je reprendrais cette théorie quand j'aborderai l'évolution.
La principale propriété qu'ont amenée les organites avec eux est la compartimentation. Ces organites ont en effet un milieu interne, de composition très différents de celle de leur milieu externe le cytoplasme et également très différents entre eux. Une autre conséquence est la multiplication des membranes. Alors que les procaryotes ne possèdent en général que la membrane plasmique, plus parfois une membrane externe, chaque organite des eucaryotes possède sont propre jeu de membranes aux propriétés très différentes les unes des autres. Tous ces éléments vont aboutir à faire de la cellule eucaryote une cellule qui bien que peu performante face à la vitalité des procaryotes, possède des potentialités absolument extraordinaires. Songez que la quasitotalité de ce que l'on observe autour de nous est constituée d'eucaryotes.
 Le noyau est l'organite qui a donné son nom aux eucaryotes (eu =
vrai, caryos = noyau), bien que quelques uns puissent en être dépourvus
à certains stades de leur existence (comme les globules rouges de
mammifères). La fonction du noyau est de contenir la majeure partie
de l'ADN cellulaire. L'ADN contenu est de type eucaryote (cette affirmation
n'est pas si ridicule que ça, comme nous le verrons avec les mitochondries).
Le noyau est l'organite qui a donné son nom aux eucaryotes (eu =
vrai, caryos = noyau), bien que quelques uns puissent en être dépourvus
à certains stades de leur existence (comme les globules rouges de
mammifères). La fonction du noyau est de contenir la majeure partie
de l'ADN cellulaire. L'ADN contenu est de type eucaryote (cette affirmation
n'est pas si ridicule que ça, comme nous le verrons avec les mitochondries).
Le noyau est limité par une enveloppe constituée d'une membrane double (deux membranes bilipidiques) qui sépare 3 compartiment, le cytoplasme à l'exterieur, le nucléoplasme à l'intérieur et le compartiment intermédiaire entre les deux membranes. Le cytoplasme et le nucléoplasme communiquent par des pores, les pores nucléaires, qui traversent les deux membranes. Au niveau des pores, la membrane interne et la membrane externe sont en continuité, il n'y a aucune communication entre le nucléoplasme et le compartiment intermédiaire (qui est en réalité une extension de la lumière du réticulum). La membrane interne de la paroi est tapissée de lamine, une protéine fibrillaire capable de fixer la chromatine.
L'intérieur du noyau abrite la chromatine, assemblage de protéines et d'ADN. Au microscope on distingue l'hétérochromatine et l'euchromatine. L'hétérochromatine, aussi appelée chromatine condensée est située généralement contre l'enveloppe et en amas isolés au centre du noyau. L'euchromatine est la forme active de l'ADN, elle apparait au microscope plus claire et plus dispersée. Une particularité des cellules des femelles de mammifères est la présence d'un condensat de chromatine près de la paroi, le corpuscule de Barr, il correspond au second chromosome X de la cellule, inactivé pour compenser l'excés par rapport aux cellules males qui n'en ont qu'un (Si les femelles avaient leurs deux chromosomes X actifs, le déséquilibre entre les gènes des deux sexes serait trop grand avec des conséquences pathologiques, comme c'est le cas quand un chromosome 21 est en trop dans la trisomie). Ce corpuscule participe au test de féminité chez les sportives.
Le noyau possède une zone spécialisée très fortement colorée par les préparations standards, le nucléole. Ce nucléole, en général unique dans les cellules, est le centre de synthèse des ARN ribosomaux et d'assemblage des sous-unités ribosomales. Ce nucléole est dynamique, il disparait avant la division cellulaire et réapparait juste après. Le nucléole est centré autour d'une structure bien particulière de l'ADN, l'organisateur nucléolaire. Il est constituée de multiples copies (plusieurs centaine de fois) des gènes de l'ARN 45S, ARN qui par clivage donne les deux grands et le petit 5,8S ARN ribosomal. Le dernier ARN ribosomal et les protéines associées sont codées ailleurs dans le génome, mais c'est quand même dans le nucléole qu'ils s'assemblent avec le reste du ribosome. L'assemblage des deux sous unités a lieu, comme chez les procaryotes, dans le cytoplasme lors de la synthèse protéique.
Pour terminer ce tout d'horizon du noyau, il faut signaler qu'à l'instar du cytoplasme qui est silloné d'un réseau fibrillaire, le cytosquelette, le noyau l'est également, par une autre famille de protéines qui constituent le nucléosquelette.
La membrane ressemble fortement à celle des procaryotes. Elle est constituée d'une double couche lipidique du même type que celle des bactéries. Elle comporte toutefois du cholestérol dans sa couche externe. Ce cholesterol sert de tampon de fluidité, il permet à la membrane de supporter de faibles écarts de température sans que cela se ressente trop dans sa consistance, un ecart de température plus important étant absorbé par un changement de longueur des lipides et de la répartition des lipides saturés et insaturé (les lipides courts ou insaturés sont plus fluides).
La membrane est traversées par des protéines : transporteurs, canaux, récepteurs aux molécules extérieures, etc... Les cellules animales comportent en plus un type protéines particulier qui traverse la membrane et fixe d'un coté le cytosquelette et de l'autre les protéines du milieu conjonctif, assurant la continuité mécanique du tissu. D'autres protéines sont absente chez les eucaryotes. Elles correspondent à des fonctions de la membrane procaryote qui sont prises en charge par des organites chez les eucaryotes. C'est le cas des protéines du complexe respiratoire qui sont localisée dans la mitochondrie.
La membrane plasmique semble en apparence plus simple que chez les procaryotes. En réalité, ce transfert de compétence vers d'autres organites à libéré la membrane pour de nouvelles fonctions. Par exemples, toutes membranes plasmique de toutes les cellules (procayotes comprises) sont polarisés, la face interne étant plus négative que la face externe. Mais les propriétés des neurones (et plus généralement des cellules excitables) proviennent de ce qu'ils sont non seulement capables de se dépolariser rapidement, mais qu'en plus ils sont capables de propager cette dépolarisation le long de leur membrane. La morphologie spécifique avec dendrites et axone n'est qu'une amélioration morphologique, les animaux les plus primitifs (éponges, cnidaires) en sont d'ailleurs dépourvus. La membrane plasmique est avec le cytosquelette l'organite qui participe le plus à la spécialisation cellulaire.
Les cellules eucaryotes sont beaucoup plus grosses que les procaryotes. Or quand le diamêtre d'une cellule augmente, sa surface augmente proportionnellement au carré et son volume au cube. Le rapport surface/volume est donc fortement diminué dans une cellule eucaryote, ce qui rend les échanges avec l'extérieur moins performants. Les eucaryotes compensent ce phénomène en se plissant, ce qui augmente la surface d'échange. Mais ces plis, s'ils sont suffisants pour fournir ses aliments à la cellule, ne permettent pas de remplir la totalité des fonctions de la membrane plasmique procaryotes. Les eucaryotes se sont alors doté d'un réseau membranaire interne le réticulum endoplasmique, qui prend en charge une partie de ces fonctions la synthèse des protéines extracellulaires et des lipides. A ces deux fonctions, s'en ajoute une troisième, particulièrement développée dans les muscles striés, le stockage du calcium intracellulaire.
Le réticulum est constituée d'un réseau de tubes ramifiés et très fortement interconnecté limité par une membrane lipidique, qui occupe la majeure partie du cytoplasme. Cette membrane est en continuité avec la membrane externe de la paroi nucléaire et l'espace du compartiment intermédiaire n'est qu'une extension de la lumière du réticulum.
L'extension du réticulum est telle qu'il peut se spécialiser
par zones. Morphologiquement, on distingue deux parties au microscope
é,lectronique, mais elles sont en réalité plus nombreuse.
Le réticulum endoplasmique granuleux porte à sa surface
une multitude de petites boules. Chacune de ces boules est un ribosome en
pleine synthèse protéique, fixé à la surface
externe du réticulum. La protéine qu'il synthétise
est injectée dans la lumière du réticulum par un pore
spécial. Dans la lumière, la protéine subira une maturation
puis elle sera envoyée à l'appareil de golgi qui l'intégrera
à une vesicule d'exocytose. Le réticulum endoplasmique granuleux
synthétise les protéines membranaires voire carrément
excrétées dans le milieu extracellulaire. La membrane nucléaire
externe fait partie de ce système.
A l'inverse le réticulum endoplasmique lisse apparait parfaitement
lisse. Ses fonctions sont très diversifiées. Il peut s'agir
tout d'abord d'une zone de réticulum granuleux mais ou ne s'exerce
pour le moment aucune synthèse. C'est aussi une zone de transit
entre les région de synthèse protéique et celles où
elles sont excrétées. Mais d'autres endroits assurent la
synthèse des lipides membranaire. Ces lipides seront intégrés
à des vésicules d'exocytose qui fourniront leurs lipides
à la membrane en fusionnant avec elle. Une autre fonction très
importante du réticulum endoplasmique lisse est de régulateur
du calcium intracellulaire. Le calcium est en effet un poison pour la plupart
des processus métaboliques, la cellule en contient donc le minimum.
Or ce calcium est utilisé comme signal par certains des récepteurs
membranaires, comme il y en a très peu dans la cellule, il suffit
de peu d'ions pour augmenter la concentration dans des proportions
élevées. Il permet entre autre de déclencher la contraction
musculaire, le potentiel d'action ou l'exocytose et même la fusion
des pronuclei lors de la fécondation. La cellule doit donc maintenir
une concentration intracellulaire de calcium très basse, tout en
s'assurant qu'il y en a assez pour le signal calcique et ensuite évacuer
le calcium du signal le plus vite possible pour permettre l'arrivée
d'un nouveau signal tout en évitant que la concentration augmente
trop et atteigne le seuil létal. Certaines zones du réticulum
lisse participent à cette régulation en constituant une reserve
de calcium pour le signal et en récupérant le calcium cytoplasmique
puis en évacuant l'excés vers le milieu extérieur.
L'appareil de Golgi travaille en relation avec le réticulum endoplasmique. C'est lui qui assure la constitution des vésicules d'exocytoses ou intracellulaires et assure leur composition correcte. L'appareil de Golgi est constitué de un ou plusieur dictyosomes. Chaque dictyosome est constitué de plusieurs saccules reliées par des pont membranaires.
Le fonctionnement du dictyosome est un peu particulier. Il est polarisé, il comporte une face de formation et une face de secrétion. La face de formation est toujour à proximité du réticulum endoplasmique lisse. Des vésicules se détachent du réticulum et fusionnent pour former une nouvelle saccule. Sur l'autre face, des vésicules se détachent et se dispersent dans le cytoplasme avant de fusionner avec leur membrane cible. Le dictyosome subit donc un renouvellement constant, une nouvelle saccule se formant toutes les 4 minutes dans une cellule en activité. C'est à l'intérieur des saccule que les protéines terminent leur maturation.
 Avec les mitochondries, on aborde un type particulier d'organites qui comprend
également les plastes végétaux et peut être
les lysosomes. Alors que les organites précédents, à
l'exception de la membrane, pouvaient être, dans certaines limites,
detruit et reconstruit par la cellule en fonction des besoins, les mitochondries
et les plastes ne peuvent pas être construit de novo à partir
de leurs éléments séparés. Une mitochondrie
provient toujours de la division d'une autre mitochondrie.
Avec les mitochondries, on aborde un type particulier d'organites qui comprend
également les plastes végétaux et peut être
les lysosomes. Alors que les organites précédents, à
l'exception de la membrane, pouvaient être, dans certaines limites,
detruit et reconstruit par la cellule en fonction des besoins, les mitochondries
et les plastes ne peuvent pas être construit de novo à partir
de leurs éléments séparés. Une mitochondrie
provient toujours de la division d'une autre mitochondrie.
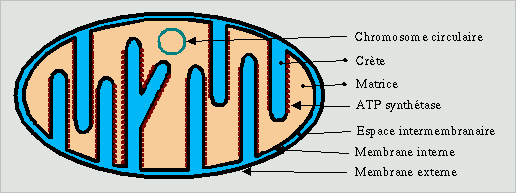
La mitochondrie est un organite de 1µm de largeur et de longueur variable. Elle est constituée de deux membranes qui isolent deux compartiments du cytoplasme. A l'intérieur de la membrane interne se trouve la matrice, la zone entre les deux membranes est appelée la zone intermembranaire.La membrane interne est repliée sur elle même et forme des crêtes alors que la membrane externe est relativement lisse. La mitochondrie est un organite mobile grace à l'action de protéines matricielles.
Les différents éléments de la mitochondrie sont de composition très différentes. La membrane externe semble n'être qu'une simple enveloppe, proche dans sa composition des membranes du réticulum endoplasmique lisse, elle s'en distingue toutefois par la présence d'une protéine particulière, la porine, qui lui assure une perméabilité totale avec la plupart des molécules cytoplasmiques. La zone intermembranaire a donc une composition peu différente de celle du cytoplasme. La membrane interne en revanche est plus interessante, elle porte toutes les protéines de la chaine réspiratoire, l'ATP synthétase et l'échangeur ATP/ADP. La matrice elle contient toutes les protéines du cycle de Krebbs. La mitochondrie peut fabriquer de l'ATP à partir du pyruvate produit par la glycolyse cytoplasmique, elle est donc capable d'exploiter la molécule de glucose au maximum de ses capacités. La mitochondrie est la centrale energétique de la cellule.
Par sa morphologie, son fonctionnement et son mode de reproduction, la mitochondrie ressemble fortement à un procaryote. Cette ressemblance a été accentuée par la découverte d'ADN dans la matrice mitochondriale. Cet ADN à une structure de type procaryote et la synthèse de protéine est assurée par des ribosomes également de type procaryotes. Ces constatations ont conduit les scientifiques à affirmer que les mitochondries seraient un ancien procaryote qui aurait été dans un lointain passé capturé par un eucaryote primitif puis intégré à sa structure. Toutefois, les protéines codées dans le chromosome ne couvrent pas l'ensemble des protéines mitochondriales, le procaryote a perdu certains gène au profit du noyau de la cellule et à également ainsi perdu toute possibilité d'autonomie. Toutes les cellules eucaryotes ont (ou ont eu) des mitochondries, la symbiose à donc du s'établir très tôt dans l'histoire à une époque ou ils étaient encore peu diversés, peut être même que c'est cet évènement qui à provoqué leur apparition. Cette théorie est dite de l'endosymbiote. Elle a été aussi proposée pour les plastes et est discutée pour d'autres organites tels que les flagelles ou les lysosomes.
Les plastes sont des organites spécifiques des végétaux. Il en existe trois types les chloroplastes, les chromoplastes et les leucoplastes qui dérivent par maturation d'un type de plaste indifférencié. L'origine des plaste semble être d'origine endosymbiotique mais leur génome est toutefois plus complet que celui des mitochondries. Comme en plus certains groupes seulement en ont acquis, la symbiose semble s'être établie plus tardivement alors que les eucaryotes étaient déjà très diversifiée. La multiplicité des mécanismes de la photosynthèse suggère en plus que la symbiose s'est produite plusieurs fois avec des symbiontes différents. Le groupe des cellules végétales qui comprend les algues et les végétaux terrestre, serait donc artificiel.
Le plus important des plaste est le chloroplaste, siège de la photosynthèse. Il est limité par une double membrane très similaire à celle des mitochondries et d'origine certainement identique. L'intérieur de l'organite est le stroma. Le stroma contient des éléments qui ressemblent à des organites intrachloroplastiques : les thylacoides qui sont des saccules allongée et des empilements de saccules plus petites, les granums, intercalés entre deux lamelles thylacoidiennes. Les chloroplastes ont donc un compartiment de plus que les mitochondries. Le stroma contient en outre l'ADN chloroplastique de type procaryote et des ribosomes de même type.
Sa fonction est de fabriquer des glucides à partir d'une source d'énergie extérieure, la lumière solaire. Il est donc en quelque sorte l'inverse de la mitochondrie qui produit de l'énergie à partir des glucides. En realité, le chloroplaste et la mitochondrie fonctionnent de façons très similaires. Tous les deux vont utiliser une source d'énergie (pyruvate pour la mitochondrie, lumière pour le chloroplaste) pour créer un gradient de proton entre l'intérieur et l'extérieur. L'energie stockée dans ce gradient permettra de synthétiser l'ATP. La grande différence est en fait dans l'utilisation de cet ATP. Alors que la mitochondrie l'exporte pour répondre aux besoin de la cellule, le chloroplaste le conserve et se sert de celui ci pour fabriquer des glucide qui seront exportés.
Le système photosynthétique est situé dans la membrane des thylacoides. Ce système à pour but de produire l'énergie nécessaire à la synthèse des glucides. Cette synthèse à lieu dans le stroma. Elle se résume en la condensation de trois molécules de CO2 en une molécule de glucide à trois carbones. La réalité est plus complexe. Ces trois CO2 vont se fixer chacun sur une molécule de glucide à cinq carbones, donnant 3 glucides à 6 carbones puis par coupure 6 glucides à trois carbones. Un des glucides sera exporté. Les cinq autres se combineront pour reformer les trois glucides de départ et permettre un nouveau cycle synthétique. Ce cycle est appellé "cycle de Benson, Bassham et Calvin" ou cycle BBC ou plus simplement cycle de Calvin.
Les glucides à trois carbones exportés par le chloroplaste seront utilisés par la cellule pour toutes les réaction où interviennent les glucides. Utilisation par les mitochondries bien sûr pour fournir l'énergie nécessaire au fonctionnement cellulaire, mais aussi mise en reserve par fabrication du glucose puis de l'amidon, synthèse du ribose et du desoxyribose constituants de l'ARN et de l'ADN, synthèse de la cellulose, etc...
La cellule eucaryote comporte d'autres organites que je vais résumer ici.

