 | | |
Le cycle de Krebs | ||
| |
Le cycle de Krebs; découvert en 1937 par le biochimiste anglais du même nom, est une chaine de réactions chimiques qui prend la suite de la glycolyse anaérobie. Toutefois, alors que la glycolyse est universellement répandue, ce cycle n'existe que chez les organismes aérobie, c'est à dire qui respirent de l'oxygène. Pour cette raison il est quelquefois improprement appelé cycle respiratoire. Ce n'est toutefois pas à ce stade qu'a lieu la véritable respiration cellulaire mais au suivant au sein de la chaine respiratoire membranaire. Son second nom de cycle des acides tricarboxylique viens de ce que deux des premiers substrats de cette chaine, le citrate et l'isocitrate sont des acides portant trois groupements COOH.
La formule globale de ce cycle est :
| CH3-CO-COOH + 3 H2O --> 3 CO2+ 10 H |
L'énergie libérée est stockée sous forme de NADH, de FADH2 et d'ATP.
Si fait le bilan avec la glycolyse qui précède, on a la formule globale :
| C6H12O6 --> 6 CO2 +12H |
Par rapport à la réaction d'oxydation totale du glucose :
|
C6H12O6 + 6 O2--> 6 CO2 + 6 H2O |
On voit que l'oxydation n'est pas complète : les hydrogène sont encore à l'état réduit. Cette étape aura lieu lors de la phase suivante la synthèse d'ATP dans la chaine respiratoire.
Le cycle de Krebs ou cycle des acides tricarboxyliques est un cycle enzymatique. Cela signifie que le premier subtrat de cette chaine est également le dernier. Ici l'oxaloacétate réagit avec l'acétyl-coenzyme A pour donner du citrate qui sera degradé par étapes successives en oxaloacétate et en CO2. L'oxaloacétate est donc disponible pour effectuer un nouveau tour du cycle avec un nouveau coenzyme A.
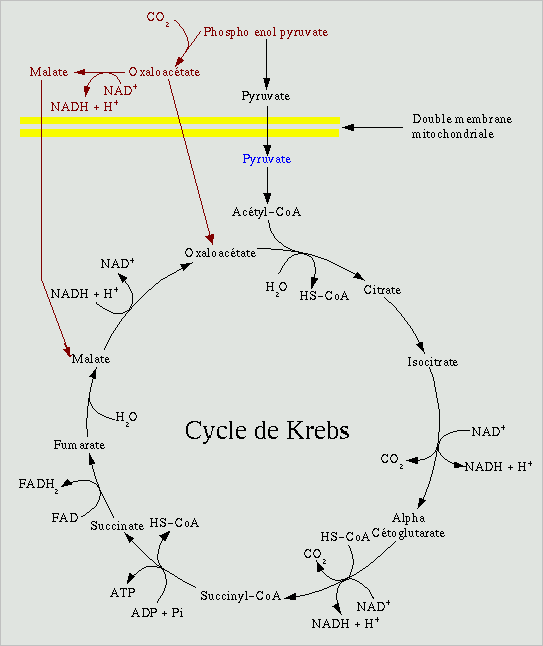 Chez les procaryotes, ce cycle se déroule dans le cytoplasme.
Chez les eucaryotes, il se déroule dans la matrice des mitochondries
(ce qui signifie que les eucaryotes sans mitochondrie ne peuvent pas respirer).
Le pyruvate produit par la glycolyse dans le cytoplasme peut toutefois pénetrer
librement dans la mitochondrie, les deux membranes lui étant totalement
perméables. Une fois dans la mitochondrie, les choses se passent
exactement comme chez les procaryotes.
Chez les procaryotes, ce cycle se déroule dans le cytoplasme.
Chez les eucaryotes, il se déroule dans la matrice des mitochondries
(ce qui signifie que les eucaryotes sans mitochondrie ne peuvent pas respirer).
Le pyruvate produit par la glycolyse dans le cytoplasme peut toutefois pénetrer
librement dans la mitochondrie, les deux membranes lui étant totalement
perméables. Une fois dans la mitochondrie, les choses se passent
exactement comme chez les procaryotes.
Le premier substrat du cycle de Krebs n'est pas le pyruvate (en bleu ci contre) mais l'acétyl coenzyme A. La première étape consiste donc a transformer le pyruvate en acétyl coenzyme A par l'intermédiaire de la pyruvate deshydrogénase, gros complexe enzymatique pouvant contenir jusqu'à soixante sous unité chez E. Coli. Au cours de cette réaction, un CO2 est dégagé et un NAD+ est réduit sous forme de NADH + H+. Le produit obtenu est formé d'un radical acétyl relié au co-enzyme A par l'intermédiaire d'une liaison thiol à haute énergie. C'est cette molécule qui va entrer dans le cycle de Krebs.
Dans un premier temps, l'acétyl coenzyme A réagit avec l'oxaloacetate pour donner du citrate, grace à la citrate synthase. L'énergie nécessaire à cette réaction est fournie par le clivage de la liaison avec le coenzyme A. Le citrate va ensuite être transformé en isocitrate par l'aconitase. L'isocitrate va être decarboxylé puis oxydé pour donner l'alpha-cétoglutarate avec réduction d'un autre NAD+ en NADH + H+. Il va à son tour subir une décarboxylation dont l'énergie p-va servir à réduire un autre NAD+ et greffer un coenzyme A à la molécule pour produire le succinyl coenzyme A, par l'action de l'alpha cétoglutarate déshydrogènase). A ce stade, tous les CO2 du pyruvate de départ ont été dégagé, mais l'énergie est encore en partie stocké dans le succinyl coenzyme A. La libération du coenzyme A pour produire le succinate va dégager de l'énergie qui va permettre de produire une molécule d'ATP à partir d'ADP et de phosphate inorganique. Le succinate va alors subir une déshydrogénation par la succinate déshydrogénase pour produire du fumarate, les deux hydrogène étant récupéré par le FAD pour donner du FADH2. Cette enzyme présente une particularité par rapport aux autres du cycle, elle est integrée à la membrane mitochondriale interne et fait partie de la chaine respiratoire. Le fumarate va être hydraté en malate par la fumarate déshydrogènase. Le malate va ensuite être oxydé en oxaloacétate avec réduction d'un autre NAD+ par la malate déshydrogénase. L'oxaloacétate, le substrat de départ est ainsi régénéré et prêt pour un nouveau cycle.
Le bilan global du cycle est donc :
| CH3-CO-COOH + 4 NAD+ + FAD + ADP + Pi + 2 H2O | |||
| ==> 3 CO2 + 4 NADH + 4 H+ + FADH2 + ATP | |||
Pour une molécule de glucose de départ, il faut multiplier ces valeurs par deux. Le cycle de Krebs produit donc 2 ATP, plus les deux de la glycolyse, cela fait 4 ATP. Les 4 NADH contienne également de l'énergie qui sera transformée en 3 ATP chacun lors de l'étape suivante (soit 24 ATP) et le FADH2 capable d'en produire 2 (4 au total). C'est donc un total de 32 ATP produit. Il ne faut pas oublier non plus les deux NADH produit lors de la glycolyse anaérobie et qui ne sont pas consommés ici (contrairement à ce qui se passe dans la fermentation). Chez les eucaryotes, ils sont produits dans le cytosol et leur entrée dans la mitochondrie consomme 1 ATP, il n'en produise que 4, soit un total final de 36 ATP, ce qui représente 37% de l'énergie stockée dans une molécule de glucose récupérée, ce qui est très nettement supérieur au rendement des fermentations (2%) . Chez les procaryotes, les réactions se produisant toutes dans le cytoplasme, le cout de l'entrée dans la mitochondrie n'existe plus et la production est de 38 ATP, le rendement est légérement supérieur (39%). Ce rendement est du même ordre de grandeur que les meilleurs appareils utilisant l'énergie chimique pour fonctionner tels les moteurs à explosion. Toutefois, l'utilisation du NADH et FADH2 produit pour fournir de l'ATP ne dépend pas du cycle de Krebs, mais d'une chaine séparée, la chaine respiratoire, dont les enzymes se situent dans l'épaisseur de la membrane interne mitochondriale.
Le cycle de Krebs est à l'intersection de nombreuses voies métaboliques. Les différents réactifs peuvent donc être soustraits du cycle. Or il ne peut fonctionner qu'à une condition, que l'oxaloacétate qui réagit avec l'acétyl coenzyme A au début soit régénéré à la fin pour refaire un nouveau cycle. Ces soustractions au cycle empechent sa régénération. Pour y remédier, des voies métaboliques (en rouge sur le schéma) permettent de produire de l'oxaloacétate et du malate à partir du phospho énol pyruvate, le substrat précédent le pyruvate dans la glycolyse. Ces deux produits peuvent ensuite traverser la double membrane mitochondriale pour relancer le cycle.
